مقدمه
چربی ها برای افزایش تراکم انرژی جیره، بهبود اجزای شیر، بهبود تولید شیر و حفظ یا بدست آوردن نمره بدنی به جیره گاوها اضافه شوند. برای اینکه یک مکمل چربی بتواند همه آن چیزها را در یک گاو شیری انجام دهد، باید چربی هضم شود. مشخص شده است که اسیدهای چرب دارای خواص فعال زیستی متفاوتی هستند و همه مکمل های چربی یا اسیدهای چرب پاسخ مشابه در گاو ندارند. به عنوان مثال، برخی از اسیدهای چرب اغلب تولید چربی شیر را افزایش می دهند در حالی که برخی دیگر برای افزایش نمره بدنی مناسب هستند. مقدار قابل توجهی از تحقیقات انجام شده است که به دلیل نقش های خاصی که اسیدهای چرب در گاو شیری بازی می کنند، درک ما را از هضم اسیدهای چرب افزایش می دهد. درک اینکه چه عواملی بر قابلیت هضم اسیدهای چرب تأثیر می گذارند به مدل ها امکان می دهد با دقت بیشتری انرژی و نتایج خاص حاصل از چربی ها را پیش بینی کنند. در حال حاضر، بیشتر نرم افزارهای تنظیم کننده جیره، اسیدهای چرب را به عنوان منبع انرژی تلقی می کنند اما لزوماً ارزش انرژی آنها را بر اساس تفاوت در ترکیب اسیدهای چرب و تفاوت در هضم اسیدهای چرب تغییر نمی دهند. ما با استفاده از اطلاعات اساسی در مورد قابلیت هضم اسیدهای چرب و همچنین مطالعات تحقیقاتی اخیر که در مورد هضم اسیدهای چرب گزارش داده اند بحث خواهیم کرد. مطالعه در این زمینه درک ما از چگونگی تفاوت هضم اسیدهای چرب و همچنین چگونگی تعامل اسیدهای چرب با یکدیگر را برای تأثیر بر قابلیت هضم افزایش داده است.
چگونه اسیدهای چرب در شکمبه تغییر می کنند
چربی که نشخوارکنندگان مصرف می کنند اغلب چربی استریفه شده غیراشباع است (تری گلیسریدها یا گلایکولیپیدها) با تنوع زیاد از مکمل های چربی تجاری در دسترس که شامل اسیدهای چرب اشباع و غیراشباع به حالت های مختلف است. شکمبه چربی های استریفه را به اسیدهای چرب و گلیسرول از طریق هیدرولیز تبدیل می کند، شکستن باند استری بین گلیسرول و اسیدهای چرب. هیدرولیز چربی ها به سرعت در شکمبه از طریق انزیم باکتری ها اتفاق می افتد (Palmquist and Jenkins, 1980). این اسیدهای چرب غیراستریفه ایجاد شده انتهای کربوکسیل ازاد دارند، که نیازمند بیوهیدروژناسیون تا اتفاق بیافتد. به این دلیل که اسیدهای چرب غیر اشباع برای گروهی خاص از میکروب ها سمی هستند، شکمبه یک روش سازگاری برای هیدروژنه کردن اسیدهای چرب غیراشباع به اسیدهای چرب اشباع گسترش داده است. اسیدهای چرب اشباع عمدتا در شکمبه خنثی هستند و بنابراین تاثیر اندکی بر تخمیر شکمبه ای دارند. هیدروژن زیادی در شکمبه به علت تخمیر غیرهوازی وجود دارد، بنابراین بیوهیدروژناسیون نسبتا سریع اتفاق می افتد. اسیدهای چربی که شکمبه را ترک می کنند اغلب اشباع هستند، برخلاف حیوانات تک معده که بخش عمده ای از مونوگلیسریدها و اسیدهای چرب غیر اشباع در مایع خروجی از معده حضور دارند. به علت این تغییرات در پروفایل اسیدهای چرب از اسیدهای چرب مصرفی به اسیدهای چرب هضم شده (یا هضم نشده)، تخمین خوراک به مدفوع اسیدهای چرب برای تخمین هضم اسیدهای چرب انفرادی در نشخوار کنندگان دقیق نیست. اگرچه در شکل چربی در شکمبه تغییراتی ایجاد می شود، اسیدهای چربی که به طور معمول در گاوهای شیری تغذیه می شوند به طور گسترده در شکمبه متابولیزه نمی شود.
هضم اسیدهای چرب در روده کوچک
مواد هضمی که شکمبه را ترک می کنند pH نزدیک خنثی دارند. در این pH، انتهای کربوکسیل ازاد اسیدهای چرب به نمک ها متصل می شوند (پتاسیم، سدیم یا کلسیم) و تشکیل نمک اسیدهای چرب را می دهند. ولیکن، pH پایین شیردان باعث جدا شدن این نمک ها از اسیدهای چرب می شود. همانطور که شیرابه هضمی به دئودنوم وارد می شود، دارای pH کمتر از 3 است (Moore and Christie, 1984). هر چربی استریفه شده که به دئودنوم می رسد سریعا هیدرولیز می شود(Doreau and Ferlay, 1994) که با یک انتهای کربوکسیل ازاد ان را ترک می کنند. ولیکن شواهدی وجود دارد که تری گلیسریدهای اشباع در مقایسه با اسیدهای چرب غیراستریفه قابلیت هضم را کاهش می دهند (de Souza and Lock, 2019) با مکمل چربی هیدروژنه شده مخصوصا قابلیت هضم را کاهش می دهند (Pantoja et al., 1996). این موضوع نشان می دهد که حتی با وجود هیدرولیز سریع در شکمبه و روده کوچک، تری گلیسیریدهای اسیدهای چرب اشباع شده ممکن است قابلیت هضم را کاهش دهند. نمک های صفراوی و فسفولیپاز لوزالمعده بر اسیدهای چرب برای تشکیل میسل هایی به منظور افزایش سطح عمل می کنند. ترشح صفرا و ترشحات لوزالمعده بدون در نظر گرفتن میزان جریان مواد هضمی نسبتاً ثابت است (Noble, 1981). نمک های صفراوی در نشخوارکنندگان غالباً تائورین مزدوج هستند که در pH پایین موثرتر هستند در مقایسه با تک معده ای ها که بیشتر گلیسین مزدوج هستند و در pH پایین موثر نیستند. در دوازدهه، pH کم و صفرا حاوی مقادیر فزاینده ای از اسید توروکولیک هستند که حلالیت اسیدهای چرب را افزایش می دهد. فسفولیپاز لوزالمعده بر لسیتین عمل کرده و لیزولسیتین، یک آمفیفیل amphiphile قوی تولید می کند. آمفیفیل ها هم خاصیت آب دوستی و هم چربی دوستی دارند و به تشکیل میسل کمک می کنند که برای انتشار اسیدهای چرب از غشا انتروسیت لازم است. لیزولسیتین یک امولسیفایر قوی برای اسید استئاریک است(Freeman 1969 and 1984) و شامل یک گروه سر قطبی و یک دم هیدروکربن است که به طور کلی به شکل لیزوفسفاتیدیل کولین است. با این حال، تغذیه 10 گرم در روز لیزولسیتین به ترتیب اثرات نسبتا کم تا منفی بر پاسخ های تولیدی هنگام تغذیه جیره با فیبر بالا و چربی غیر اشباع کم یا جیره با فیبر کم و چربی غیر اشباع بالا داشت (Rico et al., 2017a). تغذیه لسیتین سویا بدون چربی تا حدود 36/0 درصد ماده خشک تاثیری بر قابلیت هضم اسیدهای چرب 16 یا 18 کربنه نداشت (Fontoura et al., 2019; Rico et al., 2019). تغذیه لیزولسیتین غیرمحافظت شده یا لسیتین نمی تواند راه حل افزایش قابلیت هضم اسیدهای چرب در روده کوچک باشد. اسید اولئیک همچنین به صورت amphiphile برای افزایش قابلیت هضم اسید استئاریک عمل می کند(Freeman 1969 and Freeman 1984). عموما، به علت بیوهیدروژناسیون گسترده اسیدهای چرب غیراشباع میزان اسید اولئیک که به دئودنوم می رسد برای بهبود قابلیت هضم اسید استئاریک کافی نیست. ولیکن، تغذیه اسید اولئیک محافظت شده می تواند قابلیت هضم اسیدهای چرب اشباع را افزایش دهد. de Souza et al. (2018) افزایش قابلیت هضم اسیدهای چرب 16 و 18 کربنه را گزارش کردند زمانیکه مکمل چربی ترکیبی شامل 35 درصد اسید اولئیک عمدتا به صورت نمک های کلسیمی در مقایسه با ترکیب اسید پالمیتیک و اسید استئاریک تغذیه شد. نمک های کلسیمی پالم شامل 38 درصد اسید اولئیک قابلیت هضم اسیدهای چرب 16 کربنه را در مقایسه با مکمل تری گلیسرید اسید پالمیتیک افزایش می دهد (de Souza and Lock, 2018). نمک کلسیمی اسیدهای چرب حاوی اسید اولئیک به نظر می رسد که توانایی بهبود قابلیت هضم اسیدهای چرب 16 و 18 کربن را دارد، احتمالا به علت خصوصیت amphiphilic ان، که امولسیفی شدن را افزایش می دهد و به تشکیل میسل کمک می کند که برای جذب اسیدهای چرب نیاز است.
زیر مدل چربی
Moate (2004) برای توصیف قابلیت هضم روده ای اسیدهای چرب زنجیر بلند در گاوهای شیری، یک زیر مدل چربی ایجاد کرد. برای تعیین مقدار اسیدهای چرب موجود در روده باریک موجود برای هضم، این مدل مصرف اسیدهای چرب جیره، لیپولیز شکمبه ای اسیدهای چرب جیره، بیوهیدروژناسیون اسیدهای چرب زنجیره بلند، تولیدde novo اسیدهای چرب زنجیر بلند در شکمبه، عبور شکمبه ای اسیدهای چرب، و در نهایت جذب روده ای آن اسیدهای چرب را در نظر می گیرد. همه این مراحل دارای خطای ذاتی مرتبط با آنها هستند. ولیکن، اگر هدف نهایی ما براورد قابلیت هضم دقیق اسیدهای چرب است، ما نیاز داریم که براوردهای دقیقی از تمام این عوامل داشته باشیم که ترکیب و میزان اسیدهای چرب در دسترس برای جذب را تغییر می دهند. Moate (2004) گزارش کرد که رابطه خطی بین جریان روده ای اسیدهای چرب زنجیر بلند و اسیدهای چرب زنجیر بلند جذب شده وجود دارد، بطوریکه افزایش میزان اسیدهای چربی که به دوازدهه می رسند باعث جذب بیشتر اسیدهای چرب می شوند. شکل چربی همچنین در قابلیت هضم و تهیه اساس درک قابلیت هضم اسیدهای چرب انفرادی موثر است.
اندازه گیری قابلیت هضم انفرادی اسیدهای چرب
به علت تغییر در اسیدهای چرب حاصل از خوراک تا مدفوع، دامهای کانولا شده در روده برای تخمین قابلیت هضم اسیدهای چرب انفرادی نیاز است. روده بزرگ می تواند همچنین اسیدهای چرب غیراشباع را هیدروژنه کند و بیشتر جذب اسیدهای چرب در ژئوژنوم اتفاق می افتد، بنابراین نمونه های محتوای ایلئوم می تواند برای اندازه گیری قابلیت هضم اسیدهای چرب در روده کوچک مفید باشد. متناوباً، تفاوت بین اسیدهای چرب دوازدهه و مدفوع می تواند استفاده شود برای تخمین قابلیت هضم فردی اسیدهای چرب، با یک سوگیری بالقوه برعلیه قابلیت هضم اسید استئاریک، زیرا اسید چرب غیر اشباع 18 کربنی می تواند بیوهیدروژنه شود به اسید استئاریک و باعث افزایش مقدار اسید استئاریک در مدفوع شود. تعداد محدودی از مطالعات منتشر شده وجود دارد که هضم فردی اسیدهای چرب را در دوازدهه و ایلئوم گزارش می دهد. بنابراین، ما هر دو روش را ترکیب کردیم و تفاوت های ناچیز بین هضم فردی اسیدهای چرب در روش ها یافتیم. برای درک بهتر هر دو عوامل جیره ای و فاکتورهای پروفایل اسیدهای چرب که بر قابلیت هضم فردی اسیدهای چرب تأثیر می گذارند، 15 آزمایش تحقیقاتی از جمله 61 تیمار را با هم ترکیب کردیم که هضم فردی اسیدهای چرب را اندازه گیری کرد و یک متاآنالیز را برای افزایش دامنه و افزایش مشاهدات ما انجام داد (Boerman et al., 2015). همانطور که در بالا بیان شده است، به علت بیوهیدروژناسیون گسترده در شکمبه، هرچه بیشتر اسیدهای چرب غیر اشباع مصرف شوند اسیدهای چرب اشباع بیشتری در دوازدهه حضور می یابند (شکل 1). همانطور که هضم کمی از اسیدهای چرب در شکمبه اتفاق می افتد، یک رابطه مثبت بین جریان روده ای کل اسیدهای چرب و کل اسیدهای چرب مصرفی وجود دارد. زمانیکه جریان دوازدهه ای کل اسیدهای چرب افزایش می یابد، قابلیت هضم کلی اسیدهای چرب کاهش می یابد (شکل 2). ما مقایسه کردیم تمام اسیدهای چرب را با اسید استئاریک (C18:0)، زیرا اسید پالمیتیک(C16:0) یک اسید چرب اشباع است و اولئیک (C18:1)، لینولئیک (C18:2) ، و لینولنیک(C18:3) همگی اسیدهای چرب 18 کربنه غیراشباع هستند، که اسید استئاریک را یک مقایسه منطقی می سازد. متوسط هضم ظاهری روده ای بین اسید پالمیتیک و استئاریک تفاوتی ندارد. در حالی که، اسید اولئیک و لینولنیک قابلیت هضم روده ای بالاتری در مقایسه با اسید استئاریک دارند. با این حال، با همان مقایسه های مشابه برای جیره های بدون چربی مکمل، قابلیت هضم اسید استئاریک با هیچ اسید چربی متفاوت نبود و قابلیت هضم اسید استئاریک به طور قابل توجهی بیشتر از زمانی بود که مکمل های چربی در جیره گنجانده شدند. در مقادیر کمتر اسید استئاریک که به دوازدهه می رسد، قابلیت هضم اسید استئاریک زیاد است، این باعث افزایش قابلیت هضم اسید استئاریک می شود. با این حال، با افزایش جریان دوازدهه ای اسید استئاریک قابلیت هضم روده ای اسید استئاریک کاهش می یابد. کاهش قابلیت هضم اسید استئاریک با افزایش مقادیر رسیدن اسید استئاریک به دوازدهه با سایر مطالعات که کاهش هضم اسید استئاریک را در جیره های حاوی چربی در مقایسه با جیره های بدون چربی گزارش کردند مطابقت دارد (e.g. Pantoja et al., 1996). با مقدار اسیدهای چرب انفرادی رسیده به روده باریک گزارش شده در مطالعات موجود در متاآنالیز، اسید استئاریک تنها اسید چربی است که با افزایش میزان رسیدن به دوازدهه قابلیت هضم را کاهش می دهد (شکل 3). در مقایسه، به نظر می رسد اسیدهای چرب اشباع نشده دارای اثری خنثی تا مثبت است زمانیکه بیشتر به دوازدهه می رسد و قابلیت هضم اسید پالمیتیک به میزان کمتری از اسید استئاریک تغییر می کند. اشاره به این نکته مهم است که اسید استئاریک بیشترین میزان رسیدن اسیدهای چرب به دوازدهه را دارد و برخی از اسیدهای چرب اشباع نشده کمتر از ده گرم است. کاهش هضم کلی اسیدهای چرب در شکل 2 عمدتا ناشی از کاهش قابلیت هضم C18:0 است همانطور که اسیدهای چرب بیشتری به روده کوچک می رسد. یک متاآنالیز هضم اسیدهای چرب با استفاده از مشاهدات در گاو به جای استفاده از تیمارها، کاهش مشابهی را در هضم کلی اسیدهای چرب نشان داد همانطور که جریان اسیدهای چرب از شکمبه افزایش می یابد (de Souza et al., 2018). اسید استئاریک مهمترین اسید چرب خروجی از شکمبه بود و اسید چربی بود که با افزایش مقدار بیشترین کاهش هضم را داشت. قابلیت هضم اسید پالمیتیک بدون تغییر در شکمبه و خروج از شکمبه و ورود به بقیه دستگاه گوارش بود. بنابراین، مقایسه هر دو روش با استفاده از تیمارها و داده های گاو، کاهش مشابهی را در هضم اسید استئاریک نشان داد وقتی جریان دوازدهه اسید استئاریک افزایش می یابد. یک مطالعه از مکمل اسید استئاریک بسیار خالص گزارش داد که هضم کلی اسیدهای چرب هنگام افزایش مقادیر اسید استئاریک کاهش می یابد (Boerman et al., 2017). افزایش مقادیر اسید استئاریک قابلیت هضم هر دو اسید چرب 16 و 18 کربنی را کاهش می دهد، با این وجود قابلیت هضم اسید چرب 18 کربنه را بیشتر کاهش می دهد. با این حال، با افزایش مصرف اسید استئاریک، باز هم اسیدهای چرب 18 کربنی بیشتری جذب می شود. هرچه اسید استئاریک بیشتری تغذیه شود، بیشتر جذب می شود اما بازده جذب بسیار کاهش می یابد (Boerman et al. 2017). در این حالت، اگر قابلیت هضم مکمل چربی ثابت در نظر گرفته شود، به دلیل کاهش قابلیت هضم اسید چرب 18 کربنه، انرژی دریافتی گاو کاهش می یابد. یک مطالعه با مکمل اسید پالمیتیک بسیار خالص از دو جیره مختلف استفاده کرد، یکی با محتوای کل اسیدهای چرب پایین و دیگری با پنبه دانه اضافه شده و مقدار کل اسیدهای چرب بیشتر (Rico et al., 2017b).
به طور کلی، افزایش اسید پالمیتیک قابلیت هضم اسیدهای چرب 16-کربن را کاهش می دهد، اما بین جیره های با چربی کم و بالاتر تفاوت وجود دارد. جیره با چربی بالاتر کاهش قابلیت هضم کمتری از اسیدهای چرب 16 کربنی و کل داشت. این نتایج ممکن است تعجب آور باشد، زیرا به احتمال زیاد شاهد رسیدن اسید استئاریک بیشتر به دوازدهه با جیره حاوی چربی بالاتر هستیم که از تحقیقات بحث شده در بالا هضم کل اسیدهای چرب را کاهش می دهد. با این حال، چربی اضافه شده به شکل پنبه دانه ممکن است محافظت در برابر بیوهیدروژناسیون محافظت شده باشد که منجر به افزایش اسیدهای چرب غیر اشباع شده باشد که ممکن است قابلیت هضم را در مقایسه با چربی اشباع شده در دوازدهه بهبود بیشتر ببخشد. در مقایسه با مطالعه مکمل اسید استئاریک، استفاده از یک منبع کاملاً خالص اسید پالمیتیک هضم کلی اسیدهای چرب را کمتر کاهش می دهد. تفاوت بین قابلیت هضم اسیدهای چرب بر اساس محتوای اسیدهای چرب جیره پایه نشان می دهد که نه تنها مقدار اسیدهای چرب بلکه پروفایل نیز احتمالاً بر قابلیت هضم اسیدهای چرب تأثیر می گذارد.
شکل اسیدهای چرب بر قابلیت هضم تاثیر می گذارد
Daley et al. (2018) 31 مطالعه و 142 تیمار را برای هضم کلی اسیدهای چرب مختلف در 11 دسته مختلف از مکمل های چربی تجزیه و تحلیل کرد. ضرایب هضم اسیدهای چرب تحت تأثیر منبع چربی جیره است. مکمل های چربی با مقادیر بالای پالمتیک، استئاریک یا تری گلیسریدهای هیدروژنه هضم اسیدهای چرب کمتری داشتند نسبت به انهایی که مقادیر بیشتری از اسیدهای چرب اشباع نشده دارند. نتایج حاصل از Daley et al. (2018) شواهد بیشتری ارائه می دهد که فرم مکمل چربی بر قابلیت هضم تأثیر می گذارد. ما از متا رگرسیون برای تعیین اینکه چه عوامل جیره ای بر قابلیت هضم انفرادی اسیدهای چرب تأثیر می گذارند استفاده کردیم (Boerman et al., 2015). نوع چربی بر قابلیت هضم فردی اسید پالمتیک، اسید اولئیک و اسید لینولئیک تأثیر می گذارد، بطوریکه نمک های کلسیم و روغن های گیاهی قابلیت هضم عددی بالاتری دارند. به طور خاص، دانه های کامل هضم اسیدهای چرب کمتری داشتند، در حالی که دانه های کامل ممکن است تا حدی از بیوهیدروژناسیون محافظت شوند، اما هضم روده ای نیز کاهش می یابد. برای اسیدهای چرب اشباع، افزایش مصرف ماده خشک بر قابلیت هضم اسید پالمیتیک و استئاریک تأثیر منفی می گذارد. به طور بالقوه، افزایش نرخ عبور مواد خوراکی بر قابلیت هضم اسیدهای چرب اشباع به دلیل عبور خیلی سریع تأثیر منفی می گذارد تا امکان جذب در روده کوچک فراهم شود.
پروفایل اسیدهای چرب بر قابلیت هضم تاثیر می گذارد
جریان دوازدهه ای اسیدهای چرب بر قابلیت هضم فردی اسیدهای چرب تأثیر می گذارد (Boerman et al., 2015). برای اسید استئاریک، کل جریان اسیدهای چرب و همچنین نسبت اسید استئاریک و اسید اولئیک بر قابلیت هضم تأثیر منفی می گذارد. نسبت رسیدن اسید استئاریک به دوازدهه همچنین بر قابلیت هضم کل اسیدهای چرب غیر اشباع 18 کربنی تأثیر می گذارد. این نتایج نشان می دهد که پروفایلی که به دوازدهه می رسد علاوه بر مقدار فردی اسیدهای چرب، احتمالاً بر قابلیت هضم تأثیر می گذارد. به دلیل تفاوت قابلیت هضم انفرادی بین اسیدهای چرب، مقدار انرژی قابل هضم در دسترس گاو بر اساس جریان و پروفایل اسیدهای چرب متفاوت است. این تغییرات در انرژی قابل هضم بین اسیدهای چرب انفرادی تأثیر می گذارد بر تولید دام هنگامی که مکمل چربی اضافه می شود. مقدار اسیدهای چرب موجود در جیره ها در مقایسه با سایر مواد مغذی نسبتاً کم است، با این حال، از آنجا که گاوها از نظر ژنتیکی قادر به تولید شیر بیشتر هستند، ما به دنبال هر فرصتی برای افزایش تولید شیر و یا اجزای شیر هستیم. نکته مهم دیگر تأثیر مکمل های چربی بر DMI و قابلیت هضم سایر مواد مغذی است. کاهش DMI و قابلیت هضم فیبر ممکن است اثرات منفی بیشتری نسبت به کاهش قابلیت هضم اسیدهای چرب در انرژی قابل هضم داشته باشد. به منظور پیش بینی دقیق قابلیت هضم اسیدهای چرب و انرژی قابل هضم ناشی از اسیدهای چرب، ما نه تنها باید اسیدهای چرب انفرادی را در هنگام پیش بینی قابلیت هضم بررسی کنیم بلکه برهم کنش آنها با سایر اسیدهای چرب و تأثیر آنها بر قابلیت هضم سایر مواد مغذی را نیز در نظر بگیریم. گاوها مخلوط اسیدهای چرب را مصرف می کنند و به دلیل دشواری در پیش بینی میزان بیوهیدروژناسیون و میزان عبور اسیدهای چرب، دقت رسیدن اسیدهای چرب جداگانه به دوازدهه چالش برانگیز خواهد بود.
نتیجه گیری
اسید استئاریک اسید چربی است که در بیشترین مقدار به روده کوچک می رسد و برای جذب در دسترس است. افزایش میزان اسید استئاریک، قابلیت هضم اسید استئاریک و همچنین قابلیت هضم سایر اسیدهای چرب 18 کربنی را کاهش می دهد. بهبود توانایی اسیدهای چرب برای امولسیون کردن و تشکیل میسل محلی منطقی برای تلاش برای بهبود قابلیت هضم اسید استئاریک است. به نظر می رسد که اسید اولئیک محافظت شده در شکمبه بیشترین تأثیر مثبت را در افزایش قابلیت هضم اسید استئاریک دارد. شکل چربی، پروفایل اسیدهای چرب که به دوازدهه می رسد و میزان چربی همه بر قابلیت هضم انفرادی اسیدهای چرب تأثیر می گذارد. اگرچه چربی نسبتاً کمی به جیره گاوهای شیری اضافه می شود، اما حداکثر کردن میزان هضم اسیدهای چرب باعث افزایش انرژی قابل هضم و همچنین افزایش اسیدهای چرب خاص موجود برای خواص زیست فعال می شود.
References
Boerman, J. P, J. L. Firkins, N. R. St-Pierre, and A. L. Lock. 2015. Intestinal digestibility of long-chain fatty acids in lactating dairy cows: A meta-analysis and metaregression. J. Dairy Sci. 98:8889-8903.
Boerman, J. P., J. de Souza and A. L. Lock. 2017. Milk production and nutrient digestibility responses to increasing levels of stearic acid supplementation of dairy cows. J. Dairy Sci. 100:2729-2738.
Daley, V. L., L. E. Armentano, P. J. Kononoff, J. M. Prestegaard, and M. D. Hanigan. 2018. Estimation of total fatty acid content and composition of feedstuffs for dairy cattle. J. Dairy Sci. 101(Suppl. 2):295 (Abstract).
de Souza, J., C. L. Preseault, and A. L. Lock. 2018. Altering the ration of dietary palmitic, stearic, and oleic acids in diets with or without whole cottonseed effects nutrient digestibility, energy partitioning, and production responses of dairy cows. J. Dairy Sci. 101:172-185.
de Souza, J., and A. L. Lock. 2018. Short communication: Comparison of a palmitic acid-enriched triglyceride supplement and calcium salts of palm fatty acid supplement on production responses of dairy cows. J. Dairy Sci. 101:3110-3117.
de Souza, J., and A. L. Lock. 2019. Milk production and nutrient digestibility responses to triglyceride or fatty acid supplements enriched in palmitic acid. J. Dairy Sci. 102:4155-4164.
Doreau, M. and A. Ferlay. 1994. Digestion and utilization of fatty-acids by ruminants. Anim. Feed Sci. Technol. 45:379-396.
Freeman, C. P. 1984. Digestion, absorption and transport of fats - non-ruminant animals. In: Fats in Animal Nutrition. pp. 105-122. Butterworths, London, UK.
Freeman, C. P. 1969. Properties of fatty acids in dispersions of emulsified lipid and bile salt and the significance of these properties in fat absorption in the pig and the sheep. Br. J. Nutr. 23:249-263.
Fontoura, A. B. P., J. E. Rico, K. M. Keller, A. N. Davis, W. A. Myers, J. T. Siegel, R. Gervais, and J. W. McFadden. 2019. Effects of lecithin supplementation on milk production and circulating markers of metabolic health in Holstein cows. J. Dairy Sci. 102(Suppl. 1):427 (Abstract).
Moate, P. J., W. Chalupa, T. G. Jenkins, and R. C. Boston. 2004. A model to describe ruminal metabolism and intestinal absorption of long chain fatty acids. Anim. Feed Sci. & Techn. 112:79-105.
Moore, J. H. and W. W. Christie. 1984. Digestion, absorption and transport of fats in ruminant animals. In: Fats in Animal Nutrition. pp. 123-149. Butterworths, London, UK.
Noble, R. C. 1981. Digestion, transport and absorption of lipids. In W. W. Christie (Ed.) Lipid Metabolism in Ruminant Animals. pp. 57-93. Pergamon Press Ltd. Oxford, UK.
Pantoja, J., J.L. Firkins, and M.L. Eastridge. 1996. Fatty acid digestibility and lactation performance by dairy cows fed fats varying in degree of saturation. J. Dairy Sci. 79:429–437.
Palmquist, D. L., and T. C. Jenkins. 1980. Fat in lactation rations: Review. J. Dairy Sci. 62:1-14. Rico, D. E., Y. Ying, and K. J. Harvatine. 2017a. Short Communication: Effects of lysolecithin on milk fat synthesis and milk fatty acid profile of cows fed diets differing in fiber and unsaturated fatty acid concentration. J. Dairy Sci. 100:9042- 9047.
Rico J. E., J. de Souza, M. S. Allen and A. L. Lock. 2017b. Nutrient digestibility and milk production responses to increasing levels of palmitic acid supplementation vary in cows receiving diets with or without whole cottonseed. J. Anim. Sci. 95:436- 446.
Rico, J. E., A. B. P. Fontoura, B. N. Tate, and J. W. McFadden. 2019. Effects of soy lecithin on circulating choline metabolite concentrations and phosphatidylcholine profile in Holstein cows. J. Dairy Sci. 102(Suppl. 1):385 (Abstract).
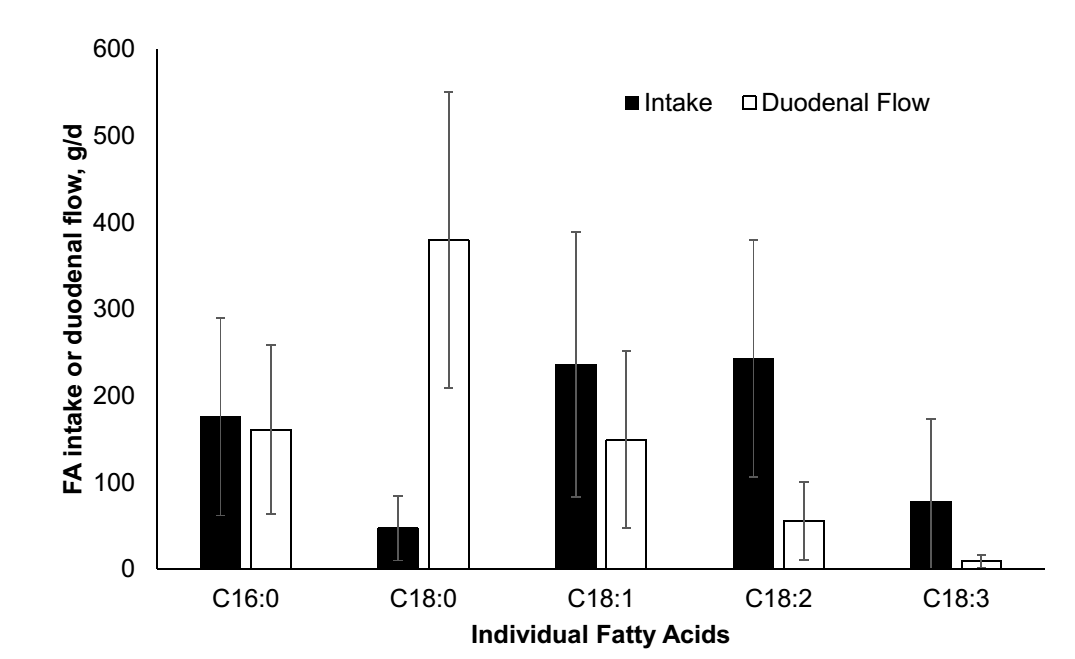
شکل 1- متوسط و انحراف استاندارد گزارش شده از 15 مطالعه بر مصرف اسیدهای چرب و جریان روده ای. به علت بیوهیدروژناسیون گسترده، کاهش در اسیدهای چرب غیراشباع رسیده به دوازدهه و افزایش در اسید استئاریک وجود دارد.
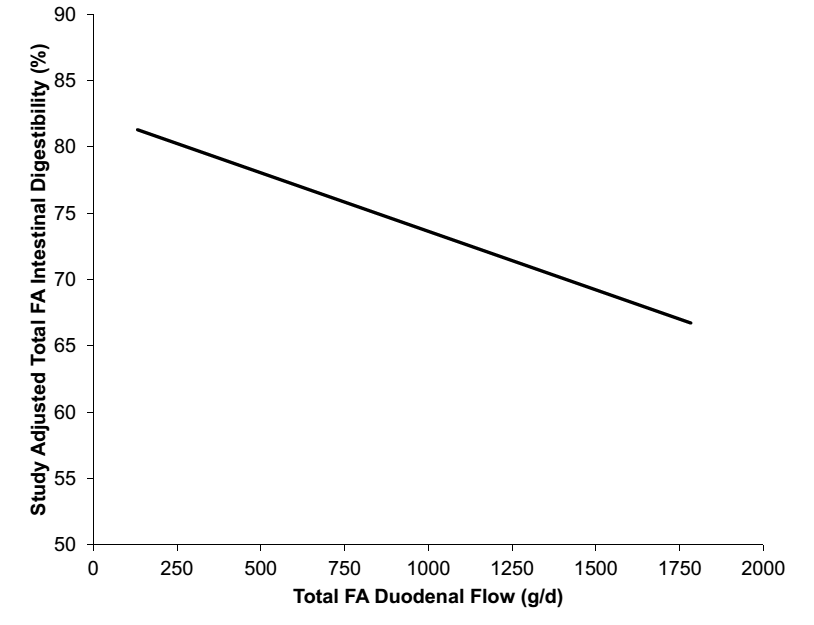
شکل 2- قابلیت هضم کل اسیدهای چرب حاصل از 61 مشاهده در 15 مطالعه که قابلیت هضم انفرادی و کلی اسیدهای چرب را گزارش کرده است که اندازه گیری شده است توسط ناپدید شده دوازده ای به ایلئوم یا مدفوع اسیدهای چرب.
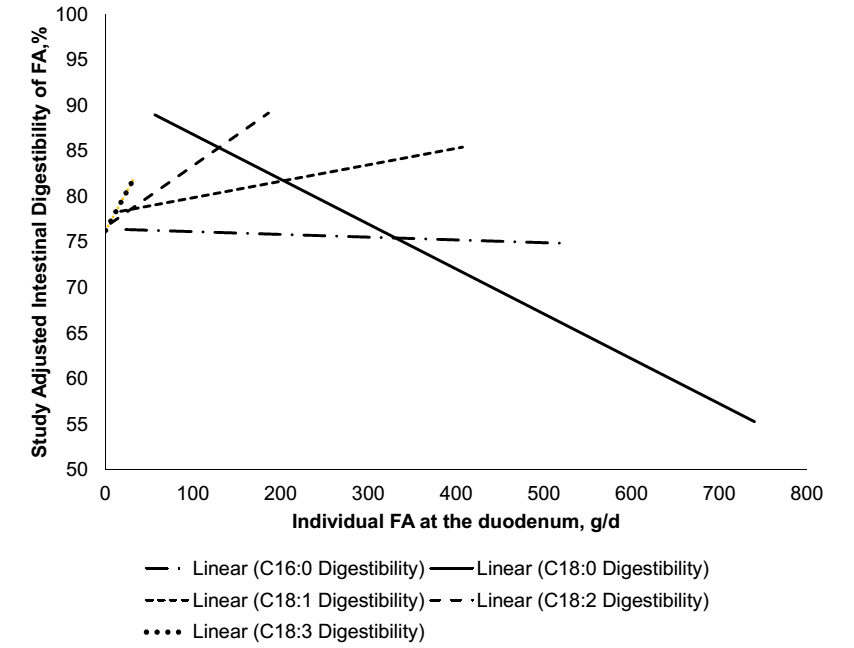
شکل 3 – مطالعات بر قابلیت هضم روده ای انفرادی اسیدهای چرب به صورت گرم از اسیدهای چرب انفرادی در روده گزارش شده در 61 تیمار از 15 مطالعه.
گروه علمی شرکت اگرین تک
